¿Qué es un pez?
Este grupo de vertebrados apareció durante la explosión cámbrica, hace unos 540 millones de años (se denomina explosión por la aparición de una gran cantidad de organismos que dio como resultado las formas animales que han llegado hasta nuestros días, además de otras muchas ya extintas), pero habrá que esperar hasta el periodo comprendido entre el Silúrico y el Devónico (de 438 a 359 millones de años de antigüedad) para que aparezcan los grupos actuales. El conjunto de estas dos últimas eras geológicas se denomina como “la edad de los peces” y fue un periodo relativamente corto en términos evolutivos teniendo en cuente la amplia diversificación acaecida, prueba del gran éxito evolutivo de este grupo.
El ancestro común de los peces y, por lo tanto, del resto de los vertebrados fue un organismo tipo anfioxo o “pez lanceta”. Se trata de un cefalocordado caracterizado por un cuerpo alargado y plano que en su estado larvario presenta notocorda. Esta estructura es equivalente a una columna vertebral y su aparición marca la transición de los invertebrados a los vertebrados.

La presencia de una estructura dorsal de naturaleza cartilaginosa permitió el desarrollo de un cordón nervioso que recorría el cuerpo del animal (origen de la médula espinal) e incorporó una novedad evolutiva notable como fue el desarrollo de un sistema sensorial sofisticado dotado de órganos complejos como los ojos. Sobre este modelo evolutivo sencillo basado en un eje longitudinal del que parten paquetes de músculos se incorporaron las aletas como elemento locomotor y se produjo el desarrollo de la región cefálica junto a la aparición de las mandíbulas, dando lugar a los primeros peces. La ventaja de estos nuevos organismos era enorme sobre otros competidores, dando lugar en el Silúrico a su diversificación y a la incorporación de sucesivas novedades adaptativas. En aquellos tiempos el clima era cálido y los casquetes polares habían liberado el agua helada creando grandes masas oceánicas epicontinentales y amplias zonas someras. Era la situación idónea para el desarrollo de la vida acuática oceánica y también continental. Los peces aprovecharon estas condiciones favorables para diversificarse, gracias en gran medida a la aparición de las mandíbulas. Estas permitían una alimentación activa y potenciaron la depredación como estrategia. Predadores y presas evolucionaron rápidamente impulsados por una carrera armamentística sin cuartel. Las nuevas técnicas de depredación generaban otras tantas para evitarla y así unos y otros fueron ocupando nuevos nichos ecológicos con una amplia variedad de especies y formas.
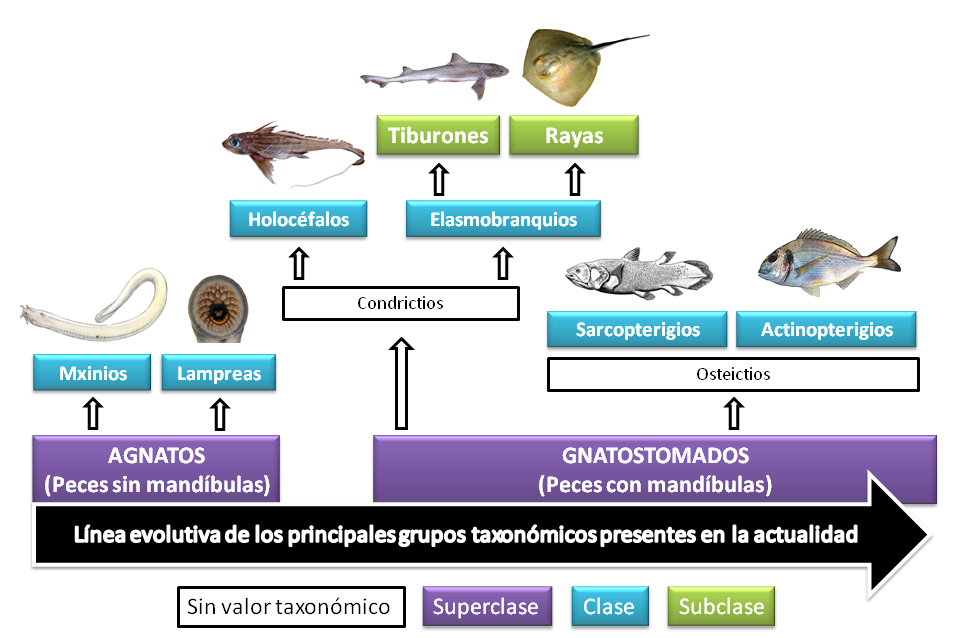
Dos hitos evolutivos marcaron la separación de los grandes grupos actuales. Por un lado, la aparición de las mandíbulas separó a los nuevos peces de los agnatos. En segundo lugar, surgió el esqueleto que diferencio a los osteíctios de los condrictios. El nivel taxonómico de estas divisiones parte de dos grandes superclases: los peces sin mandíbulas o agnatos y los peces con mandíbulas o gnatostomados. Los primeros están formados por dos clases (mixinios y lampreas) y los mandibulados por tres. Estos últimos representan el grupo más diverso y podemos hacer dos divisiones, aunque sin valor taxonómico, condrictios y osteíctios. A los condrictios corresponden dos clases: las “quimeras” (holocéfalos), cuyos arcos branquiales se abren por un solo orificio; y los tiburones (elasmobránquios), que presentan una hendidura por cada arco branquia. Los elasmobranquios se pueden dividir a su vez en dos subclases: los tiburones, con cuerpo cilíndrico e hidrodinámico; y las rayas o batoideos, que presentan un cuerpo plano con forma de disco o rombo. Por último, los osteíctios engloban otras dos clases: peces con aletas de radios duros o espinas (actinopterigios); y los peces con aletas blandas y lobuladas (sarcopterigios). Estos últimos forman una saga ancestral de los que unos pocos representantes han llegado hasta nuestros días (celacantos y peces pulmonados). Los peces pulmonados son un grupo muy importante en términos evolutivos, ya que conquistaron el medio terrestre dando lugar a la aparición de los anfibios y de todos los vertebrados terrestres.
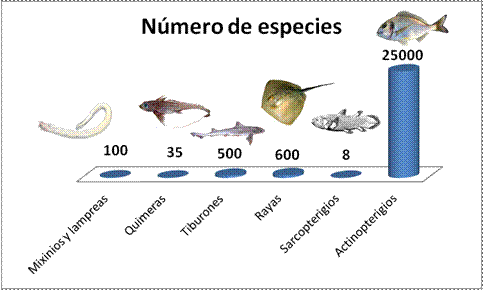
En la actualidad se calcula que existen unas 28.000 especies de “peces”, aproximadamente el 10% de todas las especies de vertebrados. El grupo mejor representado son los osteíctios y dentro de ellos los actinopterigios, con más de 26.000 especies. Los tiburones y las rayas han tenido un importante pasado, patente en un abundante registro fósil, pero su población a quedado reducida a unas 1.100 especies. Los agnatos tienen una representación bastante más discreta de algo menos de 100 especies. Por último están los peces pulmonados y los celacantos, únicamente con 8 especies conocidas. Los celacantos se creían extintos hasta que en 1938 se produjo una captura accidental en Sudáfrica. En la actualidad se han identificado dos especies de celacantos en el mundo, la sudafricana y otra en Indonesia.
MORFOLOGÍA
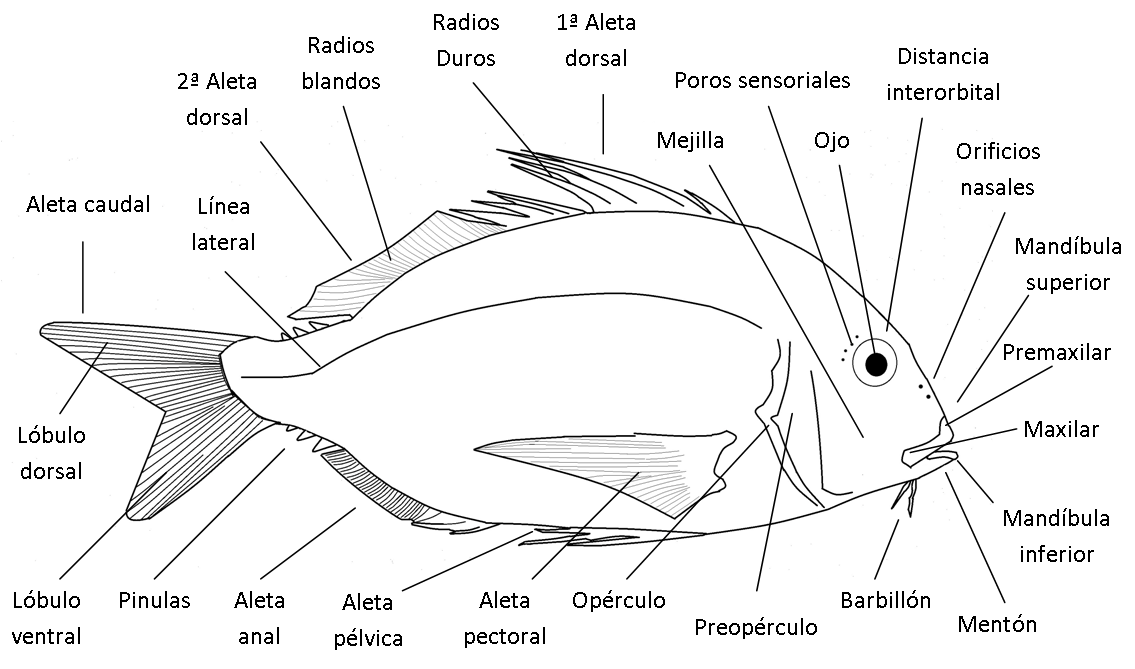
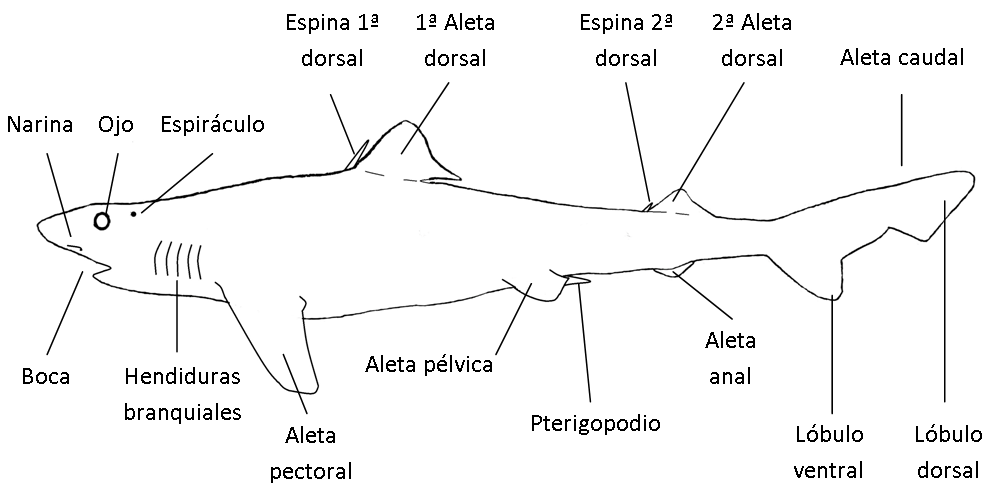
Tienen formas y tamaños muy dispares según su biología, con el cuerpo diferenciado en tres regiones: región cefálica, tronco y la región caudal.
La región cefálica abarca la zona comprendida desde la parte anterior del morro hasta la última hendidura branquial (condrictios) o hasta la parte posterior del opérculo (osteíctios). Además de la boca, los ojos, los orificios nasales o las narinas en la cabeza hay estructuras sensoriales como poros y barbillones, y otros apéndices con diferentes funciones como fotóforos. Las branquias quedan protegidas por el opérculo en los osteíctios o por un pliegue de la piel en las quimeras. En los agnatos y elasmobranquios se hayan abiertas al exterior por poros o hendiduras respectivamente.
El tronco comprende la mayor parte del cuerpo, desde el límite del opérculo hasta el inicio de la aleta anal y en él se localizan gran parte de las aletas que pueden ser pares o impares y cuya nomenclatura hacen referencia a su posición. Las aletas pectorales (APs) son pares y están situadas a cada lado del cuerpo, detrás de las aberturas branquiales. Salvo adaptaciones cponcretas no toman parte en la propulsión y se usan para maniobrar. Las segundas aletas pares suelen ser más pequeñas y son las aletas pelvianas o ventrales (APs o AVs), que pueden estar delante, detrás o al mismo nivel que las APs. Tienen funciones diversas de estabilización, defensa, sensorial o locomoción. Las aletas impares están situadas con relación al plano medio sagital del cuerpo y son las aletas dorsales (AD). Puede haber una única aleta, dos y hasta tres ADs, que se extiende longitudinalmente sobre la línea media dorsal. Su función principal es para estabilizarse durante la natación, pero puede estar modificada para cumplir funciones defensivas.
La región caudal comienza tras el inicio de la aleta anal (AA) y comprende el pedúnculo caudal y la aleta caudal (AC). El pedúnculo está dotado de una musculatura muy potente destinada a la propulsión mediante la AC. Esta aleta también hace de timón y es estabilizadora, pudiendo adoptar formas muy variadas o estar más o menos reducida. Al igual que la aleta dorsal puede estar modificada para la defensa o la caza (ejemplo de aguijones en rayas o desarrollo extremo en tiburones zorro).
Se pueden reconocer tres modelos anatómicos generales que permiten diferenciar agnatos, condrictios y osteíctios, aunque los condrictios presentan tres variaciones anatómicas.
Agnatos (ejemplo lamprea)
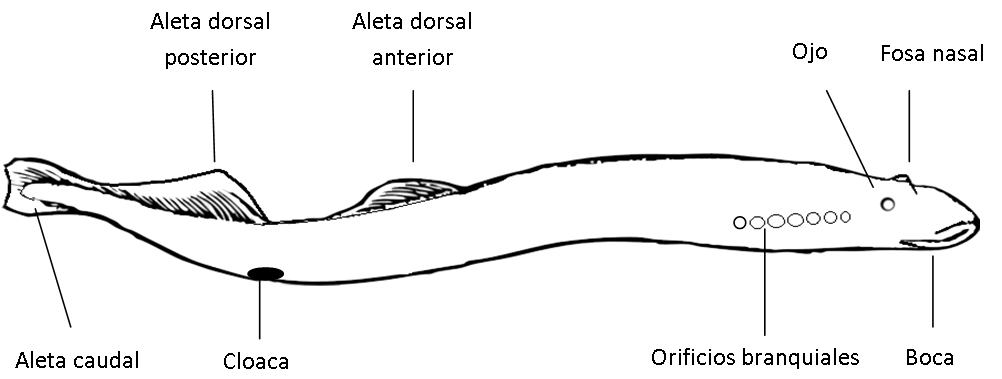
Son los más primitivos y se caracterizan por la ausencia de mandíbulas. El modelo anatómico es relativamente sencillo, sin escamas, sin aletas pares, con el cuerpo alargado y con las regiones poco diferenciadas tipo serpiente. El modelo de las lampreas presenta dos aletas dorsales y una aleta caudal. Encima de los ojos hay un orificio nasal (fosa nasal) y por detrás de éstos aparecen 7 orificios branquiales a cada lado de la cabeza. Lo más llamativo es su boca tipo ventosa y dotada con varias filas de dientes, que les permite alimentarse como hematófagos de grandes peces.
Condrictios
Los peces cartilaginosos se dividen en dos grupos con nivel de Clase: holocéfalos y elasmobranquios. Los primeros presentan cuatro pares de hendiduras branquiales cubiertas por una membrana. Mientras que los elasmobranquios tienen de cinco a siete hendiduras branquiales a cada lado de la cabeza y sin opérculo o pliegues cutáneos recubriendolas. Los holocéfalos son un pequeño grupo formado por las quimeras, mientras que los elasmobranquios son más numerosos y comprende a los tiburones propiamente dichos, los torpedos, las mantas y las rayas. Todos tienen dentículos dérmicos en lugar de escamas, pero a diferencia de los tiburones, cuya piel está completamente cubierta de dentículos dérmicos (al tacto su piel parece «papel de lija»), en las quimeras adultas la piel está desnuda y la presencia de dentículos se limita a los órganos copuladores de los machos (pterigopodios) y a un apéndice cefálico o tenáculo frontal. Los pterigopodios son otra peculiaridad de los peces cartilaginosos, que consiste en la modificación de los radios de las aletas ventrales en órganos reproductore que se emplean para la inoculación del esperma. En las quimeras se produce una calcificación de estos órganos (claspers). Respecto al resto de las aletas, los radios no son articulados y por lo tanto tienen una consistencia rígida.

Entre los condrictios hay una notable diversidad de formas con cinco modelos anatómicos distintos entre la Clases de los holocéfalos y otros cuatro dentro de la Clase de los elasmobranquios: el primer modelo corresponde al tipo quimeras (holocefalos); el segundo es la forma característicamente cilíndrica de los tiburones; por último, son tres las variaciones anatómicas de los batoideos (modelos tipo torpedos, rayas y águilas).
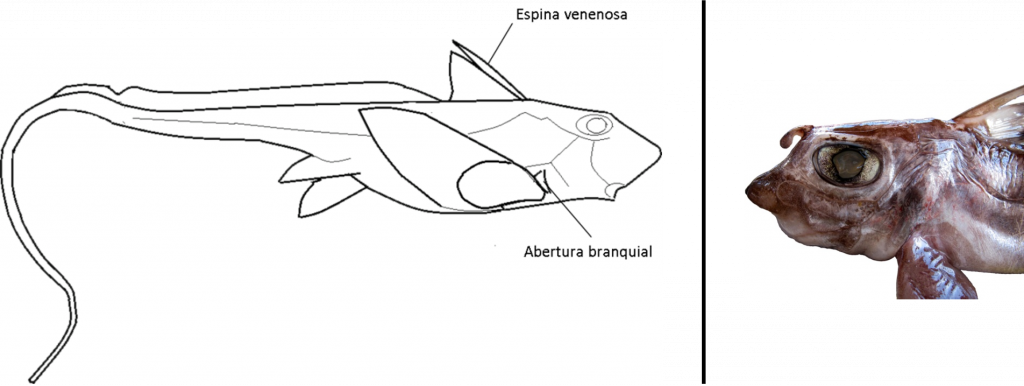
Las quimeras tienen el cuerpo alargado, algo rechoncho, con un filamento caudal largo y sin escamas. La cabeza es grande, el hocico puntiagudo y un pliegue característico cubre la cavidad branquial. El primer radio de la 1ª aleta dorsal está modificado en una espina venenosa.
El modelo fusiforme corresponde al diseño de los tiburones y está optimizado para la eficiencia hidrodinámica. El cuerpo es ancho en el centro y afilado hacia los extremos, de sección circular u ovalada. Presenta hendiduras branquiales laterales y abiertas al exterior. Las aletas son rígidas y funcionan como planos de sustentación y estabilización. El pedúnculo caudal está bien desarrollado y la aleta caudal funciona como un potente motor de avance. Este diseño es perfecto para la natación activo y la vida pelágica, donde la velocidad y la hidrodinámica son esenciales para la supervivencia.
En el modelo anatómico de los batoideos el cuerpo presenta una marcada depresión dorsoventral. Por su parte, las aletas pectorales han alcanzado un gran desarrollo, extendiéndose por su base a lo largo del tronco y la cabeza hasta fusionarse, en algunos casos, por delante del hocico. Así, cuerpo y aletas forman un conjunto con forma disco (a veces de doble disco) o romboidal. El pedúnculo caudal se elonga formando una cola gruesa y musculosa o fina tipo látigo. La aleta caudal suele estar reducida o ausente. Las aberturas branquiales se sitúan bajo las aletas pectorales, mientras que los ojos y los espiráculos están en la parte dorsal de la cabeza. Las mandíbulas presentan dientes planos adecuados para la trituración. Están adaptados mayormente a la vida sobre el lecho marino, aunque unos pocos han desarrollado hábitos pelágicos. A partir del modelo morfológico descrito se pueden diferenciar tres variaciones principales.
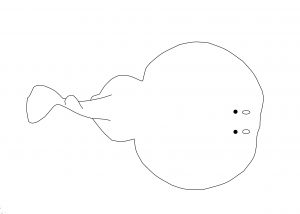
En los torpedos el conjunto cuerpo y aletas forma dos discos, con las aletas pectorales formando el disco principal y las pélvicas un disco menor. La cola es robusta, con dos aletas dorsales en la base y aleta caudal.
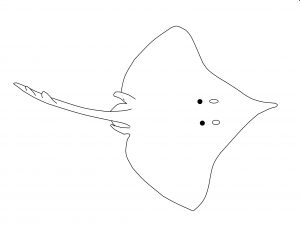
Las rayas presentan un único disco romboidal a redondeado, seguido de la cola que es más o menos robusta. Aleta caudal reducida. Es habitual la presencia de espinas dorsales.
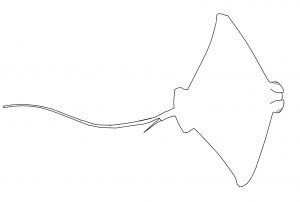
Las águilas y afines (también conocidas como chuchos) tienen un único disco romboidal de gran tamaño, seguido de una cola tipo látigo (larga y fina) con un aguijón venenoso en la base. Sin aleta caudal.
Peces óseos.
Los actinopterigios y sarcopterigios comparten un modelo anatómico similar, pero se diferencian principalmente en la estructura de las aletas. En este sentido, las aletas de los actinopterigios están formadas por un esqueleto de radios, mientras que en los sarcopterigios se trata de un apéndice carnoso (pliegue de la piel) unido al cuerpo por un solo hueso basal sobre el que se articulan otros huesos intermedios y radiales. Los radios de los actinopterigios pueden ser duros (espinas) o blandos, y estar constituidos por una varilla simple o ramificada. Otra característica de los actinopterigios es que los radios suelen estar articulados en la base, lo que permite abrir y cerrar las aletas como un abanico y plegarlas y desplegarlas con relación al cuerpo.
La piel suele estar cubierta por escamas, pero también pueden estar embutidas dentro de la piel y algunas especies carecen totalmente de ellas. En este último caso la piel está desprotegida y segregan gran cantidad de moco para protegerla. Las escamas están formadas por ganoina, que es una proteína que producen las células epidérmicas y su estructura consta de una capa de tejido óseo esponjoso y otra de tejido óseo laminar (en los peces más modernos la estructura se reduce y solo presentan el tejido óseo laminar). Se diferencian dos tipos de escamas: cicloideas y ctenoideas. Las cicloideas tiene la superficie lisa y un contorno regular, mientras que las ctenoideas suelen ser más irregulares y presentan espinas en la cara superior o sobre el borde libre (parte no imbricada de la escama).
Los peces óseos poseen un opérculo que cubre y protege las branquias. Además, su movimiento permite bombear agua sin necesidad de nadar para oxigenarse, lo cual supone un ahorro energético muy importante. Otro ahorro importante de energía se obtiene a partir de un órgano que evoluciono a partir del sistema digestivo (vejiga natatoria). Esta vejiga les permite controlar la flotación sin necesidad de moverse, llenándola o vaciándola de gases según necesiten aumentar o disminuir su flotabilidad. Los peces pulmonados carecen de vejiga natatoria, ya que la han transformado en pulmones. Éstos están conectados al esófago y les permiten respirar fuera del agua. Es un órgano homólogo a la vejiga natatoria, pero especializado en el intercambio gaseoso y no en la flotabilidad. Otra variación de este tipo de órgano se da en los celacantos, que han desarrollado un saco adiposo que, básicamente, es una vejiga vestigial llena de grasa. Esta solución les proporciona flotabilidad neutra a grandes profundidades, donde una vejiga llena de gas colapsaría por la presión ambiental.
El diseño anatómico de los actinopterigios es el modelo más exitoso y diverso entre los vertebrados acuáticos. A diferencia de los sarcopterigios, su estructura está optimizada para la ligereza, la maniobrabilidad y la velocidad en la columna de agua. Como la diversidad de formas es enorme no es posible establecer un modelo general, ya que pueden ser fusiformes, comprimidos (lateralmente), deprimidos (dorsoventralmente), planos, alargados, acintados, globosos o con especializaciones extremas que les confiere formas aberrantes. Grosso modo, el cuerpo se divide en las tres regiones referidas (región cefálica, tronco y región caudal) y las aletas pueden estar reducidas, ausentes o modificadas a órganos con diferente función (espinas venenosas, táctiles, señuelos, etc.).